Cytotoxic T Lymphocytes (CTL)
Cytotoxic T lymphocytes are lymphocytes that kill other ("target") cells.
Targets may include:
- virus-infected cells (e.g., HIV-infected CD4+ T cells);
- cells infected with intracellular bacterial or protozoal parasites;
- allografts such as transplanted kidney, heart, lungs, etc.
- cancer cells. (The tumor-infiltrating lymphocytes, TIL, that have shown some promise in cancer therapy contain CTLs.)
- In mice, and perhaps in humans, CTLs also can attack CD4+ T cells that are autoreactive; that is, T cells that recognize self antigens and thus threaten the host with autoimmunity.
- While the mechanism above protects against autoimmunity, CTLs may
trigger some autoimmune disorders, e.g. help destroy the beta cells of the islets of Langerhans, leading to Type I diabetes mellitus.
Properties of CTLs
Most of them
- belong to the CD8+ subset of T cells;
- use the αβ T-cell receptor for antigen (TCR)
- thus recognize antigens nestled in the groove of class I histocompatibility (MHC) molecules. [View]
- If they encounter (on a dendritic cell) the antigen/MHC for which their TCR is specific, they
- enter the cell cycle and go through several rounds of mitosis ("clonal expansion") followed by
- differentiation into effector ("killer") cells. Their differentiation includes forming a large number of modified lysosomes stuffed with proteins: perforin and several types of granzyme. [see below] They are helped in these activities by helper T cells that secrete stimulatory cytokines like IL-21.
- Most of these CTLs will die (of apoptosis) when they have done their job, but some (especially those that have received "help" from helper T cells) will become memory cells — long-lived cells poised to respond to the antigen if it should reappear.

An example will show the beauty and biological efficiency of CTLs.
Every time you get a virus infection, say influenza (flu), the virus invades certain cells of your body (in this case cells of the respiratory passages). Once inside, the virus subverts the metabolism of the cell to make more virus. This involves synthesizing a number of different macromolecules encoded by the viral genome.
In due course, these are assembled into a fresh crop of virus particles that leave the cell (often killing it in the process) and spread to new target cells.
Except while in transit from their old homes to their new, the viruses work inside of your cells safe from any antibodies that might be present in blood, lymph, and secretions.
But early in the process, infected cells display fragments of the viral proteins in their surface class I molecules. CTLs specific for that antigen will be able to bind to the infected cell and often will be able to destroy it before it can release a fresh crop of viruses.
In general, the role of the CD8+ T cells is to monitor all the cells of the body, ready to destroy any that express foreign antigen fragments in their class I molecules.
Some CD4+ T cells can develop into CTLs, but they can attack only those cell types (e.g. B cells, macrophages, dendritic cells) that express class II MHC molecules. Virtually every cell in the body expresses class I MHC molecules, so CD8+ CTLs are not limited in the targets they can attack.
Mechanisms of Killing
There are two.
Perforin/Granzyme Killing
CTLs have cytoplasmic granules that contain the proteins perforin and granzymes. When the CTL binds to its target, the contents of the granules are discharged by exocytosis.
- A dozen or more perforin molecules insert themselves into the plasma membrane of target cells forming a pore that enables
- granzymes to enter the cell. Granzymes are serine proteases. The two most abundant ones are
- Granzyme A. Once inside the cell, it enters the mitochondria and cleaves a subunit of complex I (the NADH dehydrogenase) of the electron transport chain producing reactive oxygen species (ROS) that kill the cell.
- Granzyme B. Once inside the cell, it proceeds to cleave the precursors of caspases thus activating them to cause the cell to self-destruct by apoptosis.
Both in structure and function, the interaction of CTL and its target resembles the synapses of the nervous system.
- The two cells are attached tightly at a small patch of plasma membrane. Special adhesion molecules hold them together.
- The granules are discharged only at that small portion of the plasma membrane (like the neurotransmitters released at a synapse).
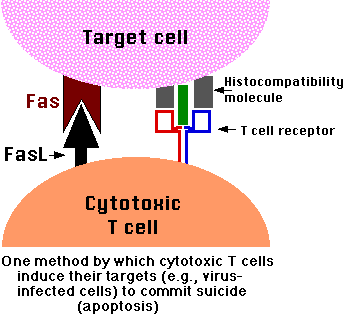
FasL/Fas Killing
CTLs express on their surface molecules of a transmembrane protein, the death activator designated Fas ligand (FasL).
Most potential CTL targets express a receptor for FasL designated Fas.
When cytotoxic T cells recognize (bind to) their target,
- they produce more FasL at their surface.
- This binds with the Fas on the surface of the target cell leading to its death by apoptosis. [Link to discussion]
Other Cytotoxic "Killer" Cells
There is another population of lymphocytes that function as "killer" cells but are not T cells. These so-called Natural Killer (NK) cells are discussed on a separate page. [Link to it.]
Unlike CTL, which are mediators of adaptive immunity; that is, learn to respond to a particular antigen, NK cells are part of the innate immune system.
4 September 2019