| Index to this page |

The cephalochordates and tunicates never develop a vertebral column. They are thus "invertebrates" and are discussed with the other invertebrates. [Link]
Although hagfishes, never replace their notochord with a vertebral column, and thus might seem not to qualify as vertebrates, they share a number of other features with other vertebrates and certainly should be classified with them. Still uncertain is whether they represent the most primitive vertebrates or are simply degenerate vertebrates (probably the latter).
All the other members of the craniata convert their notochord into a vertebral column or "backbone" (even though in some it is made of cartilage not bone). They also differ from all other animals in having quadrupled their HOX gene cluster; that is, they have 4 different clusters of HOX genes (on 4 separate chromosomes). Perhaps this acquisition played a key role in the evolutionary diversity that so characterizes the vertebrates.
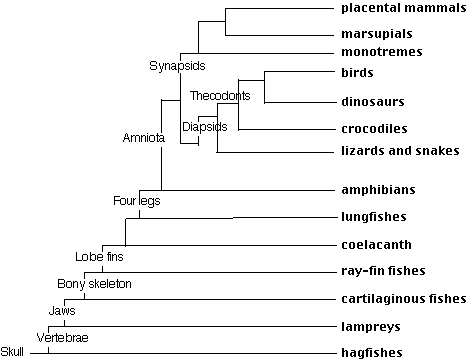
The vertebrates are subdivided into the

Lampreys and hagfishes are the only jawless vertebrates to survive today. They both have a round mouth and for this reason are often referred to as cyclostomes. They are the most primitive of the vertebrates. By "primitive", a biologist means that they are the least changed from the first vertebrates.
Besides lacking jaws,The photo (courtesy of the Carolina Biological Supply Company) is of the West Coast lamprey. Note the gill slits and the absence of paired pectoral and pelvic fins.
| Link to a discussion of the role of myelin in speeding the conduction of nerve impulses. |
With their gills exposed to sea water, all marine fishes are faced with the problem of conserving body water in a strongly hypertonic environment. Sea water is about 3.5% salt, over 3 times that of vertebrate blood. The cartilaginous fishes solve the problem by maintaining such a high concentration of urea in their blood (2.5% — far higher than the ~0.02% of other vertebrates) that it is in osmotic balance with — that is, is isotonic to — sea water.
This ability develops late in embryology, so the eggs of these species cannot simply be released in the sea. Two solutions are used:Both these solutions require internal fertilization, and the cartilaginous fishes were the first vertebrates to develop this. The pelvic fins of the male are modified for depositing sperm in the reproductive tract of the female.
Although the earliest bony fishes may have appeared late in the Silurian period, their fossils become abundant in freshwater deposits of the Devonian period. In addition to gills, these fishes had a pair of pouched outgrowths from the pharynx which served as lungs. They were inflated with air taken in through the mouth and may have provided a backup gas exchange organ when the water became too warm and stagnant to carry enough dissolved oxygen. Their kidneys were adapted for the hypotonic environment in which they lived. [Illustrated discussion]
These animals diversified through the remainder of the Devonian period (which is often called the "Age of Fishes"). Some migrated to the oceans. In this more stable environment, their lungs became transformed into a swim bladder with which they could alter buoyancy. Their kidneys became transformed as well adapting them to their new — hypertonic — surroundings. [Link to discussion]
The nostrils of bony fishes open only to the outside and are used for smelling. Some of the lobe-finned fishes developed internal openings to their nostrils. This made it possible to breath air with the mouth closed as modern lungfishes do.
Judging from present-day lungfishes, two other significant adaptations evolved in this group: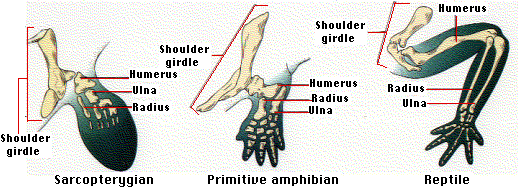
These rare modern lobe-finned fishes are the sole survivors of once-flourishing groups that also gave rise to the tetrapods — the four-legged vertebrates. In the Devonian (perhaps as early as 395 million years ago), the paired fins of some sarcopterygians moved under the body and developed limbs (complete with digits). This enabled them to venture out on land. So once again, evolution was opportunistic giving rise to the first land vertebrates, the amphibians.
The figure shows the relationship between the bones of two tetrapod forelegs and the pectoral fin of a sarcopterygian.
 With their bony limbs and lungs inherited from their lobe-finned ancestors, amphibians were so successful during the Carboniferous (Mississippian and Pennsylvanian periods) that these periods are known as the Age of Amphibians.
With their bony limbs and lungs inherited from their lobe-finned ancestors, amphibians were so successful during the Carboniferous (Mississippian and Pennsylvanian periods) that these periods are known as the Age of Amphibians.
The Carboniferous was followed by the Permian, when the earth became colder and dryer. The fortunes of the amphibians began to decline until only three groups — totaling about 8500 species — remain today:
| Link to descriptions of the heart and kidneys of amphibians. |
 The embryo developing within the egg produces 4 extraembryonic membranes: the
The embryo developing within the egg produces 4 extraembryonic membranes: the
A shelled egg is just as impervious to sperm as to water, so its arrival coincided with the development of internal fertilization.
The early aminotes soon diverged into two major lines of descent:
With the arrival of the cold, dry Permian, reptiles were well-adapted to survive because of their development of a shelled, yolk-filled egg which could be deposited on land without danger of drying out.
The photo (courtesy of the Carolina Biological Supply Company) shows an American chameleon emerging from its egg.
Other adaptations that enabled the reptiles to flourish for the next 220 million years were:This group developed the ability to convert their nitrogenous waste into uric acid. Uric acid is almost insoluble in water so its excretion involves little loss of water. (It is the whitish paste that pigeons leave on statues.) This modification largely freed the diapsids and their descendants from a dependence on drinking water; the water in their food is usually sufficient.
Diapsid evolution soon produced:
| Link to illustrated discussion of the lizard kidney |
The dominance of the reptiles during the Mesozoic era has giving it the name, the Age of Reptiles.

| View and read more about these animals |
Their amniotic egg has no shell and develops for only a brief time within the reproductive tract of the mother. During this time they receive some nourishment from the yolk sac which grows into the wall of the uterus. They are born at such an early stage of development [View] that they must crawl into a pouch on the mother's abdomen, attach themselves to one of her milk-dispensing nipples, and complete development there.
Outside of Australia, the only marsupials found today are the opossums — one species in North America, some 69 species in South America. In Australia, however, where they were protected far longer from an influx of placental mammals, there is still an broad array of marsupials. [View, for example, the wombat.]
The table shows the orders of placental mammals and representatives of each.
The members of orders shown with the same color belong to a single clade; that is, they are more closely related to each other than to the mammals in any other orders.
These clades are based on the most recent analysis of DNA sequences of several genes — both nuclear and mitochondrial — and the building of phylogenetic trees from these data.
| Tubulidentata | aardvarks |
| Macroscelidea | elephant shrews |
| Hyracoidea | hyraxes |
| Sirenia | manatee, dugong |
| Proboscidea | elephants |
| Cingulata | armadillos |
| Pilosa | anteaters, sloths |
| Dermoptera | flying lemurs |
| Primates | lemurs, New World monkeys, Old World monkeys, great apes (hominoids), humans |
| Scandentia | tree shrews |
| Rodentia | rats, mice, squirrels, beaver |
| Lagomorpha | rabbits, hares |
| Insectivora | shrews, moles |
| Chiroptera | bats |
| Pholidota | scaly anteaters |
| Carnivora | dogs, cats, lion, skunk, walrus, sea lion |
| Perissodactyla (odd-toed ungulates) |
horse, zebra, rhinoceros |
| Cetartiodactyla (even-toed ungulates and cetaceans) |
cow, sheep, pig giraffe, hippopotamus and whales, dolphins, porpoises |
The 2 orders shown in red (armadillos, sloths, giant anteaters) probably arose in the part of Gondwana that today is South America.
These 6 orders, shown in green, have been assigned to the Laurasiatheria because they are first found in the supercontinent of Laurasia. [ map and discussion ]
The even-toed ungulates and the cetaceans turn out to be so closely related that they are placed in a single order the Cetartiodactyla. In fact, hippos appear to be more closely related to the cetaceans than they are to the other even-toed ungulates.
| Return to the cladogram at the top of the page to review the evolutionary relationships of the craniata. |
| Welcome&Next Search |
{kind=link}