One of the best definitions is that of the evolutionary biologist Ernst Mayr:
A species is an actually or potentially interbreeding population that does not interbreed with other such populations when there is opportunity to do so.
Note: sometimes breeding may take place (as it can between a horse and a donkey) but if so, the offspring are not so fertile and/or well adapted as the parents (the mule produced is sterile).
The formation of two or more species often (some workers think always!) requires geographical isolation of subpopulations of the species. Only then can natural selection or perhaps genetic drift produce distinctive gene pools.
It is no accident that the various races (or "subspecies") of animals almost never occupy the same territory. Their distribution is allopatric ("other country").
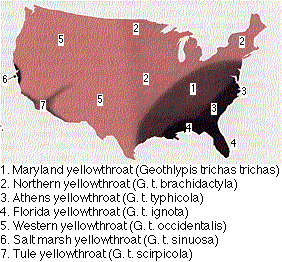
The seven distinct subspecies or races of the yellowthroat Geothlypis trichas (a warbler) in the continental U.S. would soon merge into a single homogeneous population if they occupied the same territory and bred with one another.
As a young man of 26, Charles Darwin visited the Galapagos Islands off the coast of Ecuador.
Among the animals he studied were what appeared to be 13 species* of finches found nowhere else on earth.
- Some have stout beaks for eating seeds of one size or another (#2, #3, #6).
- Others have beaks adapted for eating insects or nectar.
- One (#7) has a beak like a woodpecker's. It uses it to drill holes in wood, but lacking the long tongue of a true woodpecker, it uses a cactus spine held in its beak to dig the insect out.
- One (#12) looks more like a warbler than a finch, but its eggs, nest, and courtship behavior is like that of the other finches.
Darwin's finches. The finches numbered 1–7 are ground finches. They seek their food on the ground or in low shrubs. Those numbered 8–13 are tree finches. They live primarily on insects.
1. Large cactus finch (Geospiza conirostris)
2. Large ground finch (Geospiza magnirostris)
3. Medium ground finch (Geospiza fortis)
4. Cactus finch (Geospiza scandens)
5. Sharp-beaked ground finch (Geospiza difficilis)
6. Small ground finch (Geospiza fuliginosa)
7. Woodpecker finch (Cactospiza pallida)
8. Vegetarian tree finch (Platyspiza crassirostris)
9. Medium tree finch (Camarhynchus pauper)
10. Large tree finch (Camarhynchus psittacula)
11. Small tree finch (Camarhynchus parvulus)
12. Warbler finch (Certhidia olivacea)
13. Mangrove finch (Cactospiza heliobates)
(From BSCS, Biological Science: Molecules to Man, Houghton Mifflin Co., 1963) |
 |
| * In recent decades, teams led by Rosemary and Peter Grant have found evidence, including whole genome analysis, that several populations on different islands that were once thought to be the same species actually turn out to be different species. The current (2018) count is 20 (17 with accepted scientific names), and there is a promising candidate to be number 21 — see below.
|
Whatever the actual number turns out to be, since Darwin's time, these birds have provided a case study of how a single species reaching the Galapagos from Central or South America could — over a few million years — give rise to the various species that live there today.
Several factors have been identified that may contribute to speciation.
When the ancestor of Darwin's finches reached the Galapagos, it found
- no predators (There were no mammals and few reptiles on the islands.)
- few, if any, competitors. There were only a handful of other types of songbirds. In fact, if true warblers or woodpeckers had been present, their efficiency at exploiting their niches would surely have prevented the evolution of warblerlike and woodpeckerlike finches.
The proximity of the various islands has permitted enough migration of Darwin's finches between them to enable distinct island populations to arise. But the distances between the islands is great enough to limit interbreeding between populations on different islands. This has made possible the formation of distinctive subspecies (= races) on the various islands.
The importance of geographical isolation is illuminated by a single, fourteenth, species of Darwin's finches that lives on Cocos Island, some 500 miles (800 km) to the northeast of the Galapagos.
The first immigrants there must also have found relaxed selection pressures with few predators or competitors.
How different the outcome, though. Where one immigrant species gave rise to at least 13 on the scattered Galapagos Islands, no such divergence has occurred on the single, isolated Cocos Island.
In isolation, changes in the gene pool can occur through some combination of
These factors may produce distinct subpopulations on the different islands. So long as they remain separate (allopatric) we consider them races or subspecies. In fact, they might not be able to interbreed with other races but so long as we don't know, we assume that they could.
How much genetic change is needed to create a new species?
Perhaps not as much as you might think. For example, changes at one or just a few gene loci might do the trick. For example, a single mutation altering flower color or petal shape could immediately prevent cross-pollination between the new and the parental types (a form of prezygotic isolating mechanism).
The question of their status — subspecies or true species — is resolved if they ever do come to occupy the same territory again (become sympatric). If successful interbreeding occurs, the differences will gradually disappear, and a single population will be formed again. Speciation will not have occurred.
If, on the other hand, two subspecies reunite but fail to resume breeding, speciation has occurred and they have become separate species.
An example: The medium tree finch Camarhynchus pauper is found only on Floreana Island.
 Its close relative, the large tree finch, Camarhynchus psittacula, is found on all the central islands including Floreana.
Its close relative, the large tree finch, Camarhynchus psittacula, is found on all the central islands including Floreana.
Were it not for its presence on Floreana, both forms would be considered subspecies of the same species. Because they do coexist and maintain their separate identity on Floreana, we know that speciation has occurred.
What might keep two subpopulations from interbreeding when reunited geographically?
There are several mechanisms.
These act before fertilization occurs.
- Sexual selection — a failure to elicit mating behavior. On Floreana, Camarhynchus psittacula has a longer beak than Camarhynchus pauper, and the research teams led by Peter and Rosemary Grant have demonstrated that beak size is an important criterion by which Darwin's finches choose their mates.
- Two subpopulations may occupy different habitats in the same area and thus fail to meet at breeding time.
- In plants, a shift in the time of flowering can prevent pollination between the two subpopulations.
- Structural differences in the sex organs may become an isolating mechanism.
- The sperm may fail to reach or fuse with the egg.
These act even if fertilization does occur.
- Even if a zygote is formed, genetic differences may have become so great that the resulting hybrids are less viable or less fertile than the parental types. The sterile mule produced by mating a horse with a donkey is an example.
-
Sterility in the males produced by hybridization is more common than in females. In fact, it is the most common postzygotic isolating mechanism.
-
When Drosophila melanogaster attempts to mate with its relative Drosophila simulans, no viable males are even produced. Mutations in a single gene (encoding a component of the nuclear pore complex) are responsible.
When two species that have separated in allopatry become reunited, their prezygotic and postzygotic isolating mechanisms may become more stringent than those between the same species existing apart from each other. This phenomenon is called reinforcement. It arises from natural selection working to favor individuals that avoid interspecific matings, which would produce less-fit hybrids, when the two species are first reunited.
Hybridization between related angiosperms is sometimes followed by a doubling of the chromosome number. The resulting polyploids are now fully fertile with each other although unable to breed with either parental type. A new species has been created. This appears to have been a frequent mechanism of speciation in angiosperms.
Even without forming a polyploid, interspecific hybridization can occasionally lead to a new species of angiosperm. Two species of sunflower, the "common sunflower", Helianthus annuus, and the "prairie sunflower", H. petiolaris, grow widely over the western half of the United States. They can interbreed, but only rarely are fertile offspring produced.
However, Rieseberg and colleagues have found evidence that successful hybridization between them has happened naturally in the past. They have shown that three other species of sunflower (each growing in a habitat too harsh for either parental type) are each the product of an ancient hybridization between Helianthus annuus and H. petiolaris. Although each of these species has the same diploid number of chromosomes as the parents (2n = 34), they each have a pattern of chromosome segments that have been derived, by genetic recombination, from both the parental genomes. They demonstrated this in several ways, notably by combining RFLP analysis with the polymerase chain reaction (PCR).
They even went on to produce (at a low frequency) annuus x petiolaris hybrids in the greenhouse that mimicked the phenotypes and genotypes of the natural hybrids. (These monumental studies were described in the 29 August 2003 issue of Science.)
Another example. In Pennsylvania, hybrids between
- a species of fruit fly (not Drosophila) that feeds on blueberries and
- another species (again, not Drosophila) that feeds on snowberries
feed on honeysuckle where they neither
- encounter competition from their parental species nor
- have an opportunity to breed with them (no introgression).
(This study was published in the 28 July 2005 issue of Nature.)
In 1981, a male large cactus finch (Geospiza conirostris) turned up on Daphne Major, some 100 km (62 miles) from its home on the island of Espanola. There were no members of its species on Daphne Major, but this male mated successfully with a female medium ground finch (Geospiza fortis) living there. Their offspring then, and over the subsequent years, mated with each other but not with the resident medium ground finches (G. fortis).
Choice of a mate in these finches is highly dependent on
- body size;
- beak size;
- and song. Young birds learn their song from their father, so in this case they were imprinted on his song not that of their G. fortis mother.
Both body size and beak size were larger in the hybrids than those of the resident G. fortis finches. In addition, the larger beak of the hybrids enabled them to feed on larger and harder seeds that G. fortis did not use. So the hybrids were free from competition in their own ecological niche.
Do these hybrids represent a new species? Probably so. But until they can be shown to be unable to breed with Geospiza conirostris, we can't be sure. For the time being, the Grants are calling them "Big Bird".
So speciation can occur as a result of hybridization between two related species, if the hybrid
- receives a genome that enables it to breed with other such hybrids but
- not breed with either parental species;
- can escape to a habitat where it does not have to compete with either parent;
- is adapted to live under those new conditions.
The processes described in this page can occur over and over. In the case of Darwin's finches, they must have been repeated a number of times forming new species that gradually divided the available habitats between them. From the first arrival have come a variety of ground-feeding and tree-feeding finches as well as the warblerlike finch and the tool-using woodpeckerlike finch.
The formation of a number of diverse species from a single ancestral one is called an adaptive radiation.
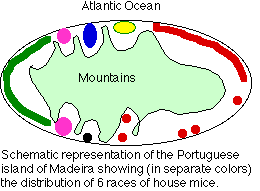 A report in the 13 January 2000 issue of Nature describes a study of house mouse populations on the island of Madeira off the Northwest coast of Africa. These workers (Janice Britton-Davidian et al) examined the karyotypes of 143 house mice (Mus musculus domesticus) from various locations along the coast of this mountainous island.
Their findings:
A report in the 13 January 2000 issue of Nature describes a study of house mouse populations on the island of Madeira off the Northwest coast of Africa. These workers (Janice Britton-Davidian et al) examined the karyotypes of 143 house mice (Mus musculus domesticus) from various locations along the coast of this mountainous island.
Their findings:
- There are 6 distinct populations (shown by different colors)
- Each of these has a distinct karyotype, with a diploid number less than the "normal" (2n = 40).
- The reduction in chromosome number has occurred through Robertsonian fusions. Mouse chromosomes tend to be acrocentric; that is, the centromere connects one long and one very short arm. Acrocentric chromosomes are at risk of translocations that fuse the long arms of two different chromosomes with the loss of the short arms.
- The different populations are allopatric; isolated in different valleys leading down to the sea.
- The distinct and uniform karyotype found in each population probably arose from genetic drift rather than natural selection.
- The 6 different populations are technically described as races because there is no opportunity for them to attempt interbreeding.
- However, they surely meet the definition of true species. While hybrids would form easily (no prezygotic isolating mechanisms), these would probably be infertile as proper synapsis and segregation of such different chromosomes would be difficult when the hybrids attempted to form gametes by meiosis.
Sympatric speciation refers to the formation of two or more descendant species from a single ancestral species all occupying the same geographic location.
Some evolutionary biologists don't believe that it ever occurs. They feel that interbreeding would soon eliminate any genetic differences that might appear.
But there is some compelling (albeit indirect) evidence that sympatric speciation can occur.
The three-spined sticklebacks, freshwater fishes that have been studied by Dolph Schluter (who received his Ph.D. for his work on Darwin's finches with Peter Grant) and his current colleagues in British Columbia, provide an intriguing example that is best explained by sympatric speciation.
They have found:
- Two different species of three-spined sticklebacks in each of five different lakes.
- a large benthic species with a large mouth that feeds on large prey in the littoral zone
- a smaller limnetic species — with a smaller mouth — that feeds on the small plankton in open water.
- DNA analysis indicates that each lake was colonized independently, presumably by a marine ancestor, after the last ice age.
- DNA analysis also shows that the two species in each lake are more closely related to each other than they are to any of the species in the other lakes.
- Nevertheless, the two species in each lake are reproductively isolated; neither mates with the other.
- However, aquarium tests showed that
- the benthic species from one lake will spawn with the benthic species from the other lakes and
- likewise the limnetic species from the different lakes will spawn with each other.
- These benthic and limnetic species even display their mating preferences when presented with sticklebacks from Japanese lakes; that is, a Canadian benthic prefers a Japanese benthic over its close limnetic cousin from its own lake.
- Their conclusion: in each lake, what began as a single population faced such competition for limited resources that
- disruptive selection — competition favoring fishes at either extreme of body size and mouth size over those nearer the mean — coupled with
- assortative mating — each size preferred mates like it
favored a divergence into two subpopulations exploiting different food in different parts of the lake.
- The fact that this pattern of speciation occurred the same way on three separate occasions suggests strongly that ecological factors in a sympatric population can cause speciation.
Sympatric speciation driven by ecological factors may also account for the extraordinary diversity of crustaceans living in the depths of Siberia's Lake Baikal.
How many genes are needed to start down the path to sympatric speciation?
Perhaps not very many.
The European corn borer, Ostrinia nubilalis, (which despite its common name is a major pest in the U.S. as well) exists as two distinct races designated Z and E. Both can be found in the same area; that is, they are sympatric.
But in the field, they practice assortative mating — only breeding with mates of their own race.
The females of both races synthesize and release a pheromone that is a sex attractant for the males. Both races use the same substance but different isomers of it. Which isomer is produced is under the control of a single enzyme-encoding gene locus. The ability of the males to respond to one isomer or the other is controlled by 2 loci.
So here is a prezygotic isolating mechanism where only 3 gene loci could begin the process of sympatric speciation. (This work is described in Lassance, J-M., et al., Nature, 22 July 2010.)
There is another possible way for new species to arise in the absence of geographical barriers.
- If a population ranges over a large area and
- if the individuals in that population can disperse over only a small portion of this range,
then gene flow across these great distances would be reduced.
The occurrence of gradual phenotypic (and genotypic) differences in a population across a large geographical area is called a cline. Successful interbreeding occurs freely at every point along the cline, but individuals at the ends of the cline may not be able to interbreed. This can be tested in the laboratory.
And, on occasions, it is tested in nature. If a cline bends around so that the ends meet, and the populations reunited at the junction cannot interbreed, then the definition of separate species has been met. Such species are called ring species and this type of speciation is called parapatric speciation.
Two examples:
1. The Caribbean slipper spurge Euphorbia tithymaloides.
Genetic analysis shows that this wildflower originated in Central America where Mexico and Guatemala share a common boundary. From there it spread in two directions
- northeast through the Yucatan peninsula and then island-hopped through Jamaica, the Dominican Republic, Puerto Rico and into the Virgin Islands;
- south through Central America, on through Venezuela, and then north through Barbados and the other islands of the Lesser Antilles finally also reaching the Virgin Islands.
Reunited in the Virgin Islands, the two populations have diverged sufficiently that they retain their distinctive genotypic and phenotypic traits. Ongoing studies will determine to what degree they may be reproductively isolated.
2. The large-blotched salamander Ensatina eschscholtzii.
This animal is found in California where it occurs in a number of different subspecies or races. A single subspecies is found in Northern California, and it is thought to be the founder of all the others. Over time that original population spread southward in two directions:
- down the Sierra Nevada mountains east of the great central San Joaquin Valley and
- down the coast range of mountains west of the valley.
South of the valley, the eastern group has moved west and now meets the western group, closing the ring. Here the two populations fail to interbreed successfully, maintaining their distinct identities. But each subspecies interbreeds in an unbroken chain up the two paths their ancestors took.
Ring species present a difficult problem in assigning species designations. It is easy to say that the populations at the ends of the cline represent separate species, but where did one give rise to the other? At every point along the cline, interbreeding goes on successfully.
The same problem faces paleontologists examining the gradual phenotypic changes seen in an unbroken line of ever-younger fossils from what one presumes to be a single line of descent. If one could resurrect the ancestral species (A) and the descendant species (B) and they could not interbreed, then they meet the definition of separate species. But there was no moment in time when one could say that A became B. So the clines of today are a model in space of Darwin's descent with modification occurring over time.
Although clines present a problem for classifiers, they are a beautiful demonstration of Darwin's conviction that the accumulation of small inherited differences can lead to the formation of new species.
31 January 2018