
This histogram shows the distribution of heights among a group of male secondary-school seniors. As you can see, the plot resembles a bell-shaped curve. Such distributions are typical of quantitative traits.
Some of the variation can be explained by differences in diet and perhaps other factors in the environment. Environment alone is not, however, sufficient to explain the full range of heights or weights.
An understanding of how genes can control quantitative traits emerged in 1908 from the work of the Swedish geneticist Nilsson-Ehle who studied quantitative traits in wheat. Using Mendel's methods, he mated pure-breeding red-kernel strains with pure-breeding white-kernel strains. The offspring were all red, but the intensity of color was much less that in the red parent. It seemed as though the effect of the red allele in the F1 generation was being modified by the presence of the white allele.

When Nilsson-Ehle mated two F1 plants, he produced an F2 generation in which red-kerneled plants outnumbered white-kerneled plants 15:1. But the red kernels were not all alike. They could quite easily be sorted into four categories. One sixteenth of them were deep red, like the P type. Four sixteenths were medium dark red, six sixteenths were medium red (like the F1 generation), and four sixteenths were light red.
The genetics of the two crosses is shown here. The alleles at one locus are indicated with prime marks; at the other, without.
These results could be explained by assuming that kernel color in wheat is controlled by not one, but two pairs of genes, the effects of which add up without distinct dominance. Each pair is located on a different chromosome or so far apart on the same chromosome that there is no linkage. Four alleles for red produce a deep red kernel. Four alleles for white produce a white kernel. Just one red allele out of four produces a light red kernel. Any two out of the four produce a medium red kernel. Any three of the four produce a medium dark red kernel.
If one plots the numbers of the different colored offspring in the F2 generation against color intensity, one gets a graph like the one below.
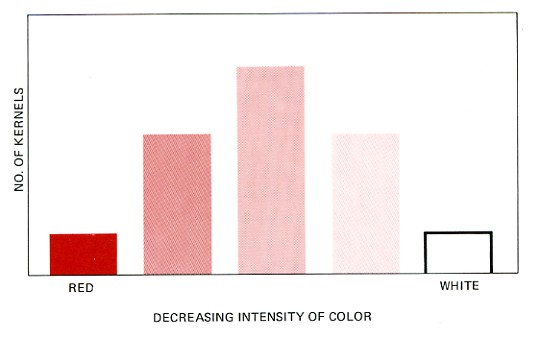
In other wheat varieties, Nilsson-Ehle found F2 generations with a ratio of red kernels to white of 63:1. These could be explained by assuming that three pairs of alleles were involved. In these cases, six different shades of red could be detected, but the color differences were very slight. Environmental influences also caused alterations in intensity so that in practice the collection of kernels displayed a continuous range of hues all the way from deep red to white.
So the occurrence of continuous variation of a trait in a population can be explained by assuming it is controlled by several pairs of genes — called quantitative trait loci (QTL) — the effects of which are added together. This is called polygenic inheritance or the multiple-factor hypothesis.
At first the study of quantitative traits was mostly confined to animal husbandry and the breeding of agricultural crops. It was based on the premise thatIn more recent times, the search for quantitative trait loci has turned to humans. A number of diseases, cancers for example, are thought to be caused by the additive effects of genes at different loci [view an example]. Pedigree analysis has provided some insights, but the use of microarrays promises to provide more.
| Welcome&Next Search |