| Index to this page |
All learning depends on memory.
The formation of memories appears to occur in two separate phases, firstHumans undergoing electroshock treatment (to alleviate their depression) are unable to remember events that occurred just prior to the treatment, but their memory of earlier events is unimpaired.
followed by formation of
Two systems that have been particularly useful in working out the cellular and molecular basis for memory formation are:
Rats and mice can be trained to solve simple tasks. For example, if a mouse is placed in a pool of murky water, it will swim about until it finds a hidden platform to climb out on. With repetition, the mouse soon learns to locate the platform more quickly. Presumably it does so with the aid of visual cues placed around the perimeter of the pool because it cannot see or smell the platform itself.
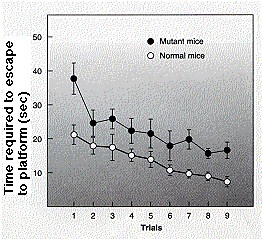
Rats or mice who have had a part of their brain called the hippocampus damaged, cannot learn this task, although they continue to solve it quickly if they were trained before their brain damage. This suggests that neurons in the hippocampus are needed for this type of learning.

The behavior of certain synapses in the "CA1" region of the hippocampus of the rat (or mouse) is consistent with their being essential for this form of long-term memory.
Slices of the hippocampus can be removed and its CA1 neurons studied in vitro with recording electrodes. Rapid, intense stimulation of presynaptic neurons evokes action potentials in the postsynaptic neuron. This is just what we would expect from the properties of synapses.
| Discussion of synapses |
What is remarkable about this system is that over time these synapses become increasingly sensitive so that a constant level of presynaptic stimulation becomes converted into a larger postsynaptic output (graph on right). This long-term potentiation can last for weeks.

Treatment of a slice of hippocampus with a drug called aminophosphonovaleric acid ("APV") blocks the formation of LTP. This is because APV blocks the action of NMDA receptors, a subset of postsynaptic receptors that normally respond to the excitatory neurotransmitter glutamate (Glu). NMDA receptors (synapse B above) are distinguished from other Glu-activated receptors in being stimulated by the drug, N-methyl-D-aspartate (NMDA).
NMDA receptors contain a transmembrane channel that allows for the facilitated diffusion of calcium ions (Ca2+) across the plasma membrane of the synapse [More]. Binding of
to these receptors opens the channel allowing Ca2+ to flow inif — and only if —
the same postsynaptic cell has been simultaneously depolarized by other synapses on it (synapse A above). (The drawing is vastly-oversimplified: each CA1 neuron has tens of thousands of synapses on it.)
The influx of Ca2+ into the neuron activates an enzyme called calcium-calmodulin-dependent kinase II (CaMKII). Kinases attach phosphate groups to proteins and, in so doing, alter their functioning. In this case, CaMKII phosphorylates a second type of Glu receptor called AMPA receptors, which makes them more permeable to sodium ions (Na+) thus lowering the resting potential of the cell and making it more sensitive to incoming impulses. In addition, there is evidence that the activity of CaMKII increases the number of AMPA receptors at the synapse.
The ability to make transgenic mice has provided tools to test this model of LTP.
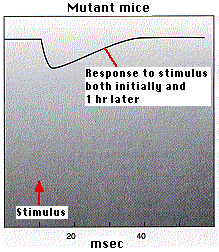
Example 1: Mice that are homozygous for a mutant CaMKII transgene fail to develop LTP.This was shown (by A. J. Silver, et. al., in Science 257:206, 1992) in two ways:
Example 2: Transgenic mice that make extra-large amounts of NMDA receptors show enhanced LTP as shown by
These findings were reported in the 2 September 1999 issue of Nature by Tang, Y-P, et al.
The experiments described above show that manipulations that affect the postsynaptic electrical response (EPSPs) of neurons stimulated electrically also affect learned behavior. They do not show that learning induces increased EPSPs in postsynaptic neurons of the hippocampus. Now, researchers at MIT have done just that.
They used rats in which they implanted an array of closely-spaced recording electrodes in the CA1 region of the hippocampus. These rats were then placed in a training apparatus where they learned — in a single trial — that moving from a lighted chamber to a dark one would give them a shock.
In just 30 minutesSo learning this conditioned response produced the electrical and synaptic changes of LTP but only in certain regions of the hippocampus. Presumably other types of learning would produce LTP in other parts of the CA1 region. (You can read the report of their work in Whitlock, J. R., et al., Science, 25 August 2006.)
Slow, weak electrical stimulation of CA1 neurons also brings about long-term changes in the synapses, in this case, a reduction in their sensitivity. This is called long-term depression or LTD. It reduces (by endocytosis) the number of AMPA receptors at the synapse. In mice, LTD results in their forgetting previous learning.
Long-term depression also occurs in
| Welcome&Next Search |
{kind=link}