
| Index to this page |

Charles Darwin and his son Francis discovered (in 1880) that the phototropic stimulus is detected at the tip of the plant.
The Darwins used grass seedlings for some of their experiments. When grass seeds germinate, the primary leaf pierces the seed coverings and the soil while protected by the coleoptile, a hollow, cylindrical sheath that surrounds it. Once the seedling has grown above the surface, the coleoptile stops growing and the primary leaf pierces it.
The Darwins found that the tip of the coleoptile was necessary for phototropism but that the bending takes place in the region below the tip.
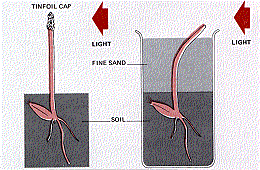
If they placed an opaque cover over the tip, phototropism failed to occur even though the rest of the coleoptile was illuminated from one side.
However, when they buried the plant in fine black sand so that only its tip was exposed, there was no interference with the tropism — the buried coleoptile bent in the direction of the light.
From these experiments, it seemed clear thatThe Danish plant physiologist Boysen-Jensen showed (in 1913) that the signal was a chemical passing down from the tip of the coleoptile.
He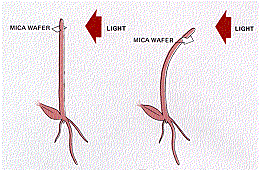 Phototropism took place normally.
Phototropism took place normally.
However, when he used a flake of impervious mica between the tip and the stump, phototropism was prevented.
Furthermore, this interference occurred only when the sheet of mica was inserted on the shady side of the preparation. When a horizontal incision was made on the illuminated side and the mica inserted in it, phototropism was normal.
This suggested that the chemical signal was a growth stimulant as the phototropic response involves faster cell elongation on the shady side than on the illuminated side.
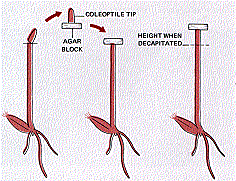
F. W. Went extracted the growth stimulant.
He removed the tips of several coleoptiles of oat, Avena sativa, seedlings. He placed these on a block of agar for several hours. At the end of this time, the agar block itself was able to initiate resumption of growth of the decapitated coleoptile. The growth was vertical because the agar block was placed completely across the stump of the coleoptile and no light reached the plant from the side.
The unknown substance that had diffused from the agar block was named auxin.
| Link to discussion of other plant responses mediated by auxin. |
The amount of auxin in coleoptile tips was far too small to be purified and analyzed chemically. Therefore, a search was made for other sources of auxin activity.
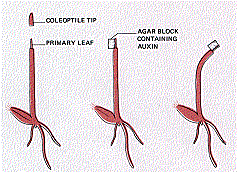
This search was aided by a technique developed by Went for determining the relative amount of auxin activity in a preparation.
The material to be assayed is incorporated into an agar block, and the block is placed on one edge of a decapitated Avena coleoptile.
As the auxin diffuses into that side of the coleoptile, it stimulates cell elongation and the coleoptile bends away from the block.
The degree of curvature, measured after 1.5 hours in the dark, is proportional to the amount of auxin activity (e.g., number of coleoptile tips used).
| Here is a photographic record of an Avena test. (Courtesy of Dr. Kenneth V. Thimann.) |
|
The use of living tissue to determine the amount of a substance is called a bioassay.
The Avena test soon revealed that substances with auxin activity occur widely in nature. One of the most potent was first isolated from human urine. It was indole-3-acetic acid (IAA) and turned out to be the auxin actually used by plants. |
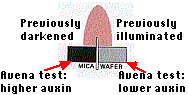
Went also discovered that it is the unequal distribution of auxin that causes the bending in phototropism. When a coleoptile tip that has previously been illuminated from one side is placed on two separated agar blocks, the block on the side that had been shaded accumulates almost twice as much auxin as the block on the previously lighted side. Hence the more rapid cell elongation on the shady side of the plant.
The growing tip of some plants tracks the movement of the sun during the day, a form of phototropism called heliotropism. At dawn, the shoot apex of the sunflower, Helianthus annuus, faces east. As the day progresses, the elongating shoot turns westward until at sunset it faces due west.
Once again, the differential accumulation of auxin in the elongating stem is responsible. In the morning, auxin levels are highest on the shady (west) side of the shoot thus bending the elongating shoot to the sunny (east) side. By sunset, auxin has redistributed to the east side of the shoot bending the shoot to the west. During the night, auxin gradually redistributes back to the west side while the shoot gradually reorients toward the coming sunrise.
Heliotropism maximizes the exposure of the growing tip to the light needed for photosynthesis. In the case of the sunflower, it also improves the efficiency of pollination.
Gravitropism also involves the unequal distribution of auxin.
 When an oat coleoptile tip is placed on two separated agar blocks, as shown here, there is no difference in the auxin activity picked up by the two blocks. When the preparation is placed on its side, however, the lower block accumulates twice as much auxin activity as the upper block. Under natural conditions, this would cause greater cell elongation on the underside of the coleoptile and the plant would bend upward.
When an oat coleoptile tip is placed on two separated agar blocks, as shown here, there is no difference in the auxin activity picked up by the two blocks. When the preparation is placed on its side, however, the lower block accumulates twice as much auxin activity as the upper block. Under natural conditions, this would cause greater cell elongation on the underside of the coleoptile and the plant would bend upward.
 of roots.
of roots.
The graph (based on the work of K. V. Thimann) shows the effect of auxin concentration on root and stem growth. The difference between the behavior of roots and stems lies in the difference in the sensitivity of their cells to auxin. Auxin concentrations high enough to stimulate stem growth inhibit root growth.
| Welcome&Next Search |