
Symbiotic nitrogen fixation occurs in plants that harbor nitrogen-fixing bacteria within their tissues. The best-studied example is the association between legumes and bacteria in the genus Rhizobium.
Each of these is able to survive independently (soil nitrates must then be available to the legume), but life together is clearly beneficial to both. Only together can nitrogen fixation take place.A symbiotic relationship in which both partners benefits is called mutualism.
| Link to discussion of the role of nitrogen fixation in the nitrogen cycle. |
Rhizobia are Gram-negative bacilli that live freely in the soil (especially where legumes have been grown). However, they cannot fix atmospheric nitrogen until they have invaded the roots of the appropriate legume.
The interaction between a particular strain of rhizobia and the "appropriate" legume is mediated by:
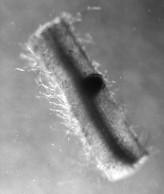
If the combination is correct, the bacteria enter an epithelial cell of the root; then migrate into the cortex. [Link to root structure.] Their path runs within an intracellular channel that grows through one cortex cell after another. This infection thread is constructed by the root cells, not the bacteria, and is formed only in response to the infection.
When the infection thread reaches a cell deep in the cortex, it bursts and the rhizobia are engulfed by endocytosis into membrane-enclosed symbiosomes within the cytoplasm. At this time the cell goes through several rounds of mitosis — without cytokinesis — so the cell becomes polyploid.
The electron micrograph on the right (courtesy of Dr. D. C. Jordan) shows a rhizobia-filled infection thread growing into the cell (from the upper left to the lower right). Note how the wall of the infection thread is continuous with the wall of the cell. The dark ovals are the symbiosomes.

The cortex cells then begin to divide rapidly forming a nodule. This response is driven by the translocation of cytokinins from epidermal cells to the cells of the cortex.
The photo on the left (courtesy of The Nitragin Co. Milwaukee, Wisconsin) shows nodules on the roots of the birdsfoot trefoil, a legume.

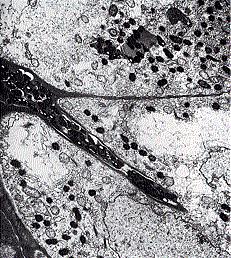
The rhizobia also go through a period of rapid multiplication within the nodule cells. Then they begin to change shape and lose their motility. The bacteroids, as they are now called, may almost fill the cell. Only now does nitrogen fixation begin.
The electron micrograph on the right (courtesy of R. R. Hebert) shows bacteroid-filled cells from a soybean nodule. The horizontal line marks the walls between two adjacent nodule cells.
Root nodules are not simply structureless masses of cells. Each becomes connected by the xylem and phloem to the vascular system of the plant. The photo below on the left shows a developing lateral root on a pea root. On its right is a segment of a pea root showing a developing nodule 12 days after the root was infected with rhizobia. Both structures are connected to the nutrient transport system of the plant (dark area extending through the center of the root). (Photomicrographs courtesy of the late John G. Torrey.)
Thus the development of nodules, while dependent on rhizobia, is a well-coordinated developmental process of the plant.
| |
Although some soil bacteria (e.g., Azotobacter) can fix nitrogen by themselves, rhizobia cannot. Clearly rhizobia and legumes are mutually dependent.
The benefit to the legume host is clear. The rhizobia make it independent of soil nitrogen.
But why is the legume necessary? The legume is certainly helpful in that it supplies nutrients to the bacteroids with which they synthesize the large amounts of ATP needed to convert nitrogen (N2) into ammonia (NH3)
In addition, the legume host supplies one critical component of nitrogenase — the key enzyme for fixing nitrogen.
The bacteroids need oxygen to make their ATP (by cellular respiration). However, nitrogenase is strongly inhibited by oxygen. Thus the bacteroids must walk a fine line between too much and too little oxygen. Their job is made easier by another contribution from their host: hemoglobin.
Nodules are filled with hemoglobin. So much of it, in fact, that a freshly-cut nodule is red. The hemoglobin of the legume (called leghemoglobin), like the hemoglobin of vertebrates, probably supplies just the right concentration of oxygen to the bacteroids to satisfy their conflicting requirements.

The metal molybdenum is a critical component of nitrogenase and so is absolutely essential for nitrogen fixation. But the amounts required are remarkably small. One ounce (28.3 g) of molybdenum broadcast over an acre (0.4 hectare) of cropland in Australia was found to be sufficient to restore fertility for over ten years.
The photo at the right shows that the legume clover grows normally only where the supply of molybdenum is adequate. The soil shown here (in eastern Australia) is naturally deficient in molybdenum. Although the entire fenced-in plot was seeded to clover, the plant was able to flourish and fix nitrogen only where molybdenum fertilizer had been added (foreground). (Photo courtesy of A. J. Anderson.)Because of the specificity of the interaction between the Nod factor and the receptor on the legume, some strains of rhizobia will infect only peas, some only clover, some only alfalfa, etc. The treating of legume seeds with the proper strain of rhizobia is a routine agricultural practice. (The Nitragin Company, that supplied one of the photos above specializes in producing rhizobial strains appropriate to each leguminous crop.)
Although symbiotic nitrogen fixation is most common among the legumes, it also occurs sporadically in other plant groups. Notable examples are alders (found in temperate regions around the world), bayberries and sweetfern (found in northeastern U.S.), and casuarina trees (an Australian native now found in many semitropical seashores). In all these plants, the symbiont in the nodules is not rhizobia, but Frankia, an actinobacterium.
The distribution of symbiotic nitrogen fixation in all these groups is "patchy", i.e., some species have it (perhaps acquired by independent acquisition of the trait — convergent evolution) while some of their very close relatives do not (perhaps they lost the trait as they evolved from ancestors that had it. If so, why they abandoned what seems to be such a beneficial trait is a mystery).
There are many examples of symbiotic nitrogen fixation in surface ocean waters. In each case, the host is a unicellular diatom. The symbiont is a cyanobacterium in some cases, a rhizobium in others.
| Welcome&Next Search |